 Nghiên cứu của chúng tôi tập trung vào sự hiểu biết về các cơ chế phân tử mà các đột biến trong rhodopsin và trong các gen điều chỉnh hoạt động buôn bán của nó, dẫn đến các bệnh về võng mạc. Rhodopsin là một thành phần chính của màng cảm nhận ánh sáng của tế bào cảm thụ ánh sáng hình que võng mạc, nơi nó có chức năng như một GPCR cảm nhận ánh sáng, bắt đầu dòng truyền dẫn quang. Rhodopsin cũng là chất điều hòa trung tâm sức khỏe của tế bào cảm thụ ánh sáng và có thể gây ra hơn 150 đột biến khác nhau gây ra bệnh viêm võng mạc sắc tố. Các đột biến ảnh hưởng đến đầu tận cùng của rhodopsin gây ra các dạng nghiêm trọng của bệnh viêm võng mạc sắc tố chiếm ưu thế trên thể nhiễm sắc (ADRP). Trong quá trình nghiên cứu của mình, chúng tôi đã xác định mô-típ VxPx ở đầu cuối C rhodopsin là một tín hiệu nhắm mục tiêu theo đường mật được bảo tồn (CTS). Chúng tôi đã xác định các thành phần của phức hợp nhắm mục tiêu thể mật nhận ra tín hiệu này để điều chỉnh hoạt động buôn bán rhodopsin và đổi mới màng tế bào cảm thụ hình que võng mạc.
Nghiên cứu của chúng tôi tập trung vào sự hiểu biết về các cơ chế phân tử mà các đột biến trong rhodopsin và trong các gen điều chỉnh hoạt động buôn bán của nó, dẫn đến các bệnh về võng mạc. Rhodopsin là một thành phần chính của màng cảm nhận ánh sáng của tế bào cảm thụ ánh sáng hình que võng mạc, nơi nó có chức năng như một GPCR cảm nhận ánh sáng, bắt đầu dòng truyền dẫn quang. Rhodopsin cũng là chất điều hòa trung tâm sức khỏe của tế bào cảm thụ ánh sáng và có thể gây ra hơn 150 đột biến khác nhau gây ra bệnh viêm võng mạc sắc tố. Các đột biến ảnh hưởng đến đầu tận cùng của rhodopsin gây ra các dạng nghiêm trọng của bệnh viêm võng mạc sắc tố chiếm ưu thế trên thể nhiễm sắc (ADRP). Trong quá trình nghiên cứu của mình, chúng tôi đã xác định mô-típ VxPx ở đầu cuối C rhodopsin là một tín hiệu nhắm mục tiêu theo đường mật được bảo tồn (CTS). Chúng tôi đã xác định các thành phần của phức hợp nhắm mục tiêu thể mật nhận ra tín hiệu này để điều chỉnh hoạt động buôn bán rhodopsin và đổi mới màng tế bào cảm thụ hình que võng mạc.
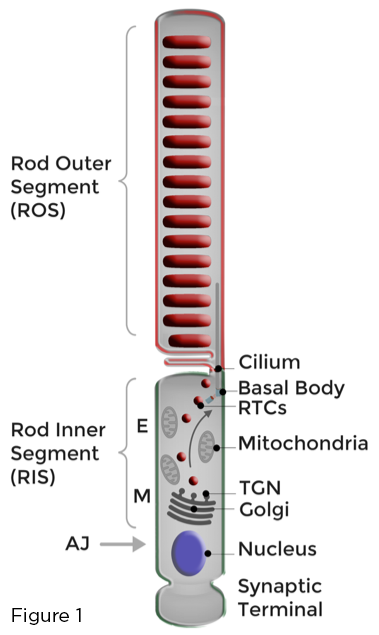 Nền tảng cho chức năng tối ưu của tế bào cảm quang hình que là sự ngăn chặn nghiêm ngặt của màng cảm quang chứa nhiều rhodopsin thành cilium sơ cấp được biến đổi độc đáo tạo thành phân đoạn ngoài hình que (ROS). Cân bằng nội môi của cơ quan thụ cảm quang hình que được duy trì bằng cách bổ sung liên tục các màng ROS bị tổn thương do ánh sáng và hấp thụ hiệu quả các protein và lipid ROS tham gia vào quá trình truyền quang, tránh xa các protein tham gia vào các chức năng tế bào đa dạng diễn ra trong cơ thể tế bào lân cận có tên là phân đoạn bên trong hình que (RIS), hoặc ở đầu cuối synap hình que. Golgi và các bào quan sinh tổng hợp khác được bản địa hóa trong RIS, trong vùng myoid (M). Hoạt động buôn bán màng phân cực trong các thanh võng mạc liên quan đến việc tổng hợp, phân loại và vận chuyển, thông qua RIS, một lượng lớn chất mang rhodopsin hướng Golgi-to-cilia (RTC). Các RTC đi ngang qua vùng ellipsoid (E) chứa đầy ti thể và hợp nhất với màng sinh chất RIS ở gần cilium. Các màng mới được tổng hợp sau đó được chuyển đến ROS (Hình 1).
Nền tảng cho chức năng tối ưu của tế bào cảm quang hình que là sự ngăn chặn nghiêm ngặt của màng cảm quang chứa nhiều rhodopsin thành cilium sơ cấp được biến đổi độc đáo tạo thành phân đoạn ngoài hình que (ROS). Cân bằng nội môi của cơ quan thụ cảm quang hình que được duy trì bằng cách bổ sung liên tục các màng ROS bị tổn thương do ánh sáng và hấp thụ hiệu quả các protein và lipid ROS tham gia vào quá trình truyền quang, tránh xa các protein tham gia vào các chức năng tế bào đa dạng diễn ra trong cơ thể tế bào lân cận có tên là phân đoạn bên trong hình que (RIS), hoặc ở đầu cuối synap hình que. Golgi và các bào quan sinh tổng hợp khác được bản địa hóa trong RIS, trong vùng myoid (M). Hoạt động buôn bán màng phân cực trong các thanh võng mạc liên quan đến việc tổng hợp, phân loại và vận chuyển, thông qua RIS, một lượng lớn chất mang rhodopsin hướng Golgi-to-cilia (RTC). Các RTC đi ngang qua vùng ellipsoid (E) chứa đầy ti thể và hợp nhất với màng sinh chất RIS ở gần cilium. Các màng mới được tổng hợp sau đó được chuyển đến ROS (Hình 1).
Việc phân loại thành các RTC được điều chỉnh bởi các GTPases nhỏ của họ Rab và Arf, đóng vai trò trung tâm trong việc tổ chức vận chuyển màng nội bào cũng như phân phối màng đến các lông mao sơ cấp. Các protein liên quan đến quá trình hình thành và duy trì lông mao được mã hóa bởi khoảng 25% gen bệnh võng mạc di truyền, với các đột biến gây thoái hóa võng mạc, thận có nang, béo phì và khuyết tật ống thần kinh trong một loạt các rối loạn di truyền, được gọi chung là bệnh ciliopathies.
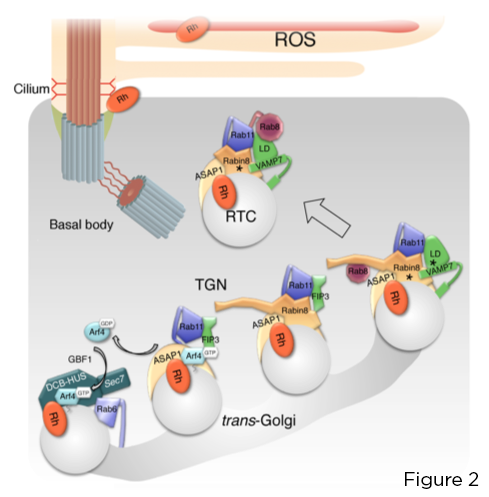 GTPase Arf4 nhỏ nhận dạng và liên kết trực tiếp với VxPx CTS đầu cuối rhodopsin C. Arf4, được kích hoạt tại cơ quan thụ cảm ánh sáng Golgi bởi yếu tố trao đổi nucleotide Arf guanine (GEF) GBF1, bắt đầu lắp ráp theo từng bước mối quan hệ nhắm mục tiêu tập trung vào protein kích hoạt Arf GTPase (GAP) ASAP1, làm trung gian thủy phân GTP trên Arf4 và Rab11a -Phức hợp hiệu ứng kép FIP3-Rabin8. Phức hợp này kiểm soát việc lắp ráp mô-đun nhắm mục tiêu Rab11a-Rabin8-Rab8 được bảo tồn cao, trực tiếp tuyển dụng R-SNARE VAMP7 vào các RTC để điều chỉnh sự hợp nhất của chúng tại cơ sở mật, thông qua ghép nối VAMP7 với màng sinh chất cognate SNAREs cú pháp 3 và SNAP-25 (Hình 2).
GTPase Arf4 nhỏ nhận dạng và liên kết trực tiếp với VxPx CTS đầu cuối rhodopsin C. Arf4, được kích hoạt tại cơ quan thụ cảm ánh sáng Golgi bởi yếu tố trao đổi nucleotide Arf guanine (GEF) GBF1, bắt đầu lắp ráp theo từng bước mối quan hệ nhắm mục tiêu tập trung vào protein kích hoạt Arf GTPase (GAP) ASAP1, làm trung gian thủy phân GTP trên Arf4 và Rab11a -Phức hợp hiệu ứng kép FIP3-Rabin8. Phức hợp này kiểm soát việc lắp ráp mô-đun nhắm mục tiêu Rab11a-Rabin8-Rab8 được bảo tồn cao, trực tiếp tuyển dụng R-SNARE VAMP7 vào các RTC để điều chỉnh sự hợp nhất của chúng tại cơ sở mật, thông qua ghép nối VAMP7 với màng sinh chất cognate SNAREs cú pháp 3 và SNAP-25 (Hình 2).
Điểm mấu chốt của thác nước mật Rab11a-Rabin8-Rab8 là Rab8 GEF Rabin8, một protein giàn giáo đa chức năng tương tác với các protein thể mật chọn lọc, chẳng hạn như phức hợp vận chuyển TRAPPII và BBSome, cho thấy vai trò trung tâm trong đường dẫn mật của các thụ thể cảm giác. Chức năng của nó bị ảnh hưởng bởi các đột biến ở NDR2 kinase (STK38L), được mã hóa bởi gen thoái hóa võng mạc sớm ở chó (erd) tương ứng với bệnh ciliopathy Leber bẩm sinh ở người (LCA). Nghiên cứu hiện tại của chúng tôi cho thấy GFP-Rabin8 của con người biểu hiện trong quá trình chuyển gen X. laevis tạo màu với Rabin8 và rhodopsin nội sinh tại Golgi và trên RTC, mở đường cho các nghiên cứu trong tương lai về vai trò của Rabin8 trong sự tiến triển của màng dọc theo đường mật, vốn có khả năng bị gián đoạn trong các bệnh thoái hóa võng mạc di truyền.
Nói chung, các nghiên cứu của chúng tôi tiết lộ rằng nhắm mục tiêu màng tới ROS là một dạng nhắm mục tiêu theo đường mật được bảo tồn. Mô típ VxPx có trong các protein màng mật khác. Nhắm mục tiêu phức hợp dựa trên Arf4 được bảo tồn nhắm mục tiêu các thụ thể cảm giác đến các lông mao chính thông qua các mạng lưới chức năng phức tạp của các GTPase nhỏ và các cơ quan điều hòa của chúng rất nhạy cảm với các đột biến gây thoái hóa võng mạc và các tế bào liên kết.
Deretic D., Lorentzen E và Fresquez T. (2019). Phần trong và ngoài của phức hợp nhắm mục tiêu màng mật dựa trên Arf4. GTPases nhỏ. GTPases nhỏ. Ngày 2019 tháng 9 năm 1: 12-10.1080. doi: 21541248.2019.1616355 / XNUMX. [Epub trước khi in]
Kandachar V, Tam BM, Moritz OL và Deretic D. (2018) Một mạng tương tác giữa SNARE VAMP7 và Rab-GTPases trong một phức hợp nhắm mục tiêu màng thể mật. Tế bào J. (2018) J Cell Sci. 2018 Tháng mười hai 10; 131 (24). pii: jcs222034. doi: 10.1242 / jcs.222034.
Wang J., Fresquez T. Kandachar V. và Deretic D (2017). Arf GEF GBF1 và Arf4 hiệp lực với hàng hóa thụ thể cảm giác, rhodopsin, để điều chỉnh vận chuyển qua màng mật. Tế bào J. J Cell Sci. 2017 Ngày 1 tháng 130; 23 (3975): 3987-10.1242. doi: 205492 / jcs.2017. Epub 12 ngày XNUMX tháng XNUMX.
Vetter M, Wang J, Lorentzen E, và Deretic D. (2015) Địa hình mới lạ của mạng tương tác hiệu ứng Rab11 trong một phức hợp nhắm mục tiêu màng thể mật. GTPases nhỏ. 2015 Ngày 2 tháng 6; 4 (165): 73-10.1080. doi: 21541248.2015.1091539 / 2015. Epub 23 ngày XNUMX tháng XNUMX.
Wang J, và Deretic D. (2015) Arf và Rab11 effector FIP3 hoạt động hiệp đồng với Arf GAP ASAP1 để hướng Rabin8 nhắm mục tiêu thụ thể mật. Tế bào J. J Cell Sci jcs.162925; Advance Online Article ngày 11 tháng 2015 năm 10.1242, doi: 162925 / jcs.XNUMX (2015).
Wang J. và Deretic D (2014). Các phức hợp phân tử chỉ đạo vận chuyển rhodopsin đến các lông mao sơ cấp. Prog Retin Eye Res. 2014 tháng 38; 1: 19-10.1016. doi: 2013.08.004 / j.preteyeres.2013. Epub 14 ngày XNUMX tháng XNUMX
Wang J, Morita Y, Mazelova J và Deretic D. (2012). Arf GAP ASAP1 cung cấp một nền tảng để điều chỉnh việc nhắm mục tiêu thụ thể qua trung gian Arf4-và Rab11-Rab8. EMBO J 31, 4057-4071.
Mazelova J, Ransom N, Astuto-Gribble L, Wilson MC và Deretic D. (2009) Ghép cặp Syntaxin 3 và SNAP-25, được điều chỉnh bởi axit docosahexaenoic omega-3 (DHA), kiểm soát việc phân phối rhodopsin cho quá trình sinh học của lông mao các bào quan cảm giác có nguồn gốc, các đoạn ngoài hình que. J. Tế bào khoa học. 122, 2003-2013.
Mazelova J, Astuto-Gribble L, Inoue H, Tam BM, Schonteich E, Prekeris R, Moritz OL, Randazzo PA và Deretic D. (2009) Mô típ nhắm mục tiêu theo đường mật VxPx chỉ đạo việc lắp ráp mô-đun buôn bán thông qua Arf4. EMBO J. 28, 183-192.
Deretic D, Williams AH, Ransom N, Morel V, Hargrave PA và Arendt A. (2005) Rhodopsin C-endinus, vị trí đột biến gây bệnh võng mạc, điều chỉnh quá trình buôn bán bằng cách liên kết với ARF4. Proc. Natl. Học viện Khoa học HOA KỲ. 102: 3301-3306.